HT 2 – Ökologie, Evolution
Thema: Das Schweigen der Grillen
1.
Gib eine Definition für den Begriff der reproduktiven Isolation an und ermittle einen möglichen Isolationsmechanismus zwischen den in Material A genannten Grillenarten (Material A). Erläutere die Bedeutung von Lock- und Balzgesang der Grillenmännchen unter evolutiven Aspekten (Material A).
(12 Punkte)
2.
Fasse die in den Abbildungen 2 und 3 dargestellten Ergebnisse zusammen (Material B) und leite Selektionsvorteile und Selektionsnachteile für Teleogryllus oceanicus-Männchen ab (Materialien A und B). Gib die Unterschiede zwischen konvergenten und divergenten Entwicklungen an und analysiere vor diesem Hintergrund umfassend Abbildung 4 (Materialien A bis C).
(28 Punkte)
3.
Erkläre die zu erwartenden Zahlenverhältnisse für die angegebenen Erbgänge (Tabelle 1) und werte die tatsächlichen Kreuzungsergebnisse im Hinblick auf den Erbgang aus (Material D).
(10 Punkte)
4.
Erkläre die Entwicklung der stummen Grillen auf Kauai mithilfe der Synthetischen Evolutionstheorie (Materialien A und B). Erläutere die Geschwindigkeit des evolutionären Wandels bei Teleogryllus oceanicus (Materialien A bis D).
(16 Punkte)
Material A: Gesang der Feldgrillen
In Australien kommen zwei Arten von Feldgrillen vor, die morphologisch kaum zu unterscheiden sind. Beide Arten sind dunkelbraun bis schwarz und etwa drei Zentimeter groß. Die Verbreitungsgebiete von Teleogryllus oceanicus und Teleogryllus commodus überlappen sich im Osten von Australien. Wie viele andere Feldgrillen erzeugen die Männchen mithilfe ihrer Vorderflügel einen Gesang. Dazu bewegen sie die Schrillader, eine harte, gezähnte Struktur auf der Unterseite des rechten Vorderflügels, über die Hinterkante des linken Vorderflügels hin und her. Den Grillenweibchen fehlt die gezähnte Schrillader. Die von den Männchen erzeugten Laute können als Gesangsmuster aufgezeichnet und verglichen werden (Abbildung 1). Der Lockgesang der Grillenmännchen lockt paarungsbereite Weibchen an, dann nehmen die Partner über die Fühler Kontakt auf. Während des weiteren Paarungsrituals spielen chemische Signale und ein Balzgesang des Männchens eine wichtige Rolle. Der Balzgesang ist energieaufwändiger als der Lockgesang und innerhalb der jeweiligen Grillenart variabel. Weibchen bevorzugen Männchen mit hochfrequenten, energieaufwändigen Balzgesängen. Der Lockgesang verschiedener Männchen innerhalb einer Art ist hingegen sehr ähnlich.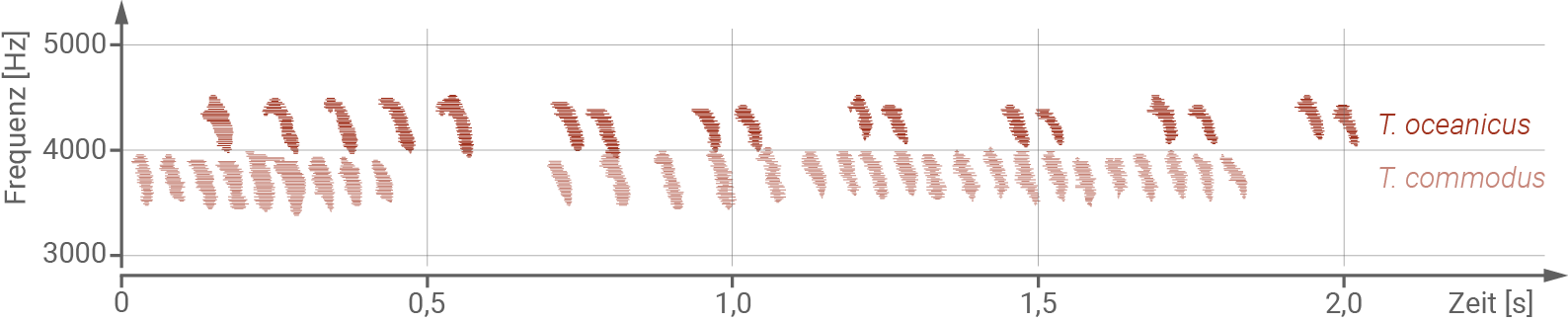
Abb. 1: Gesangsmuster des Lockgesangs der Feldgrillen.
Die Frequenz in Hertz (Hz, Schwingungen pro Sekunde) ist ein Maß für die Tonhöhe.
Die Frequenz in Hertz (Hz, Schwingungen pro Sekunde) ist ein Maß für die Tonhöhe.
Material B: Teleogryllus oceanicus auf der Hawaii-Inselgruppe
Während der Besiedlung der hawaiianischen Inseln durch den Menschen vor etwa tausend Jahren wurde T. oceanicus vermutlich von Australien zur Hawaii-Inselgruppe verschleppt und Populationen etablierten sich auf den verschiedenen Inseln wie Hawaii, Kauai und Oahu. Eine ursprünglich in Nordamerika heimische Fliege, Ormia ochracea, die etwa 1989 auf die HawaiiInselgruppe eingeschleppt worden ist, kann die Lockgesänge der Männchen von T. oceanicus hören. Die Weibchen von O. ochracea lokalisieren rufende Grillen-Männchen und legen ihre Larven auf oder in direkter Nähe der Grillenmännchen ab. Die Fliegenlarven bohren sich in das Innere der Grillenmännchen, entwickeln sich dort weiter und fressen dabei die Grille auf. Nach etwa sieben Tagen ist die Entwicklung der Fliegen vollendet. Bei ihrer Freisetzung stirbt das Grillenmännchen. Auf Kauai waren 1991 rund 30 Prozent der rufenden T. oceanicus-Männchen von den Fliegen befallen. 2001 konnten nur noch wenige Grillen dieser Art auf Kauai gefunden werden, darunter nur ein rufendes Männchen. Im Jahr 2003 wurde wieder eine größere Population von T. oceanicus entdeckt, deren Männchen eine deutlich veränderte Morphologie der Flügel zeigten. Unter anderem war bei ihnen die Schrillader stark verkürzt. Diese Männchen können weder Lockgesang noch Balzgesang produzieren. Die stummen T. oceanicus-Männchen von Kauai werden als K-flatwing-Männchen bezeichnet. Innerhalb von weniger als 20 Generationen hatte sich auf Kauai eine stabile K-flatwing-Population etabliert.Zwei Jahre später wurden auf Oahu, einer Nachbarinsel von Kauai, ebenfalls stumme Männchen entdeckt, die als O-flatwing-Männchen bezeichnet werden. Innerhalb weniger Generationen etablierte sich eine stabile O-flatwing-Population auf Oahu (Abbildung 2). Von vielen anderen Feldgrillen-Arten ist bekannt, dass Männchen, die gerade keinen Lockgesang produzieren, sich häufig in räumlicher Nähe zu rufenden Feldgrillen-Männchen aufhalten. Bei Freilanduntersuchungen auf Hawaii, Oahu und Kauai wurde der Abstand von T. oceanicus-Männchen zu einem Lautsprecher gemessen, der den T. oceanicus-Lockgesang abspielte (Abbildung 3 ).
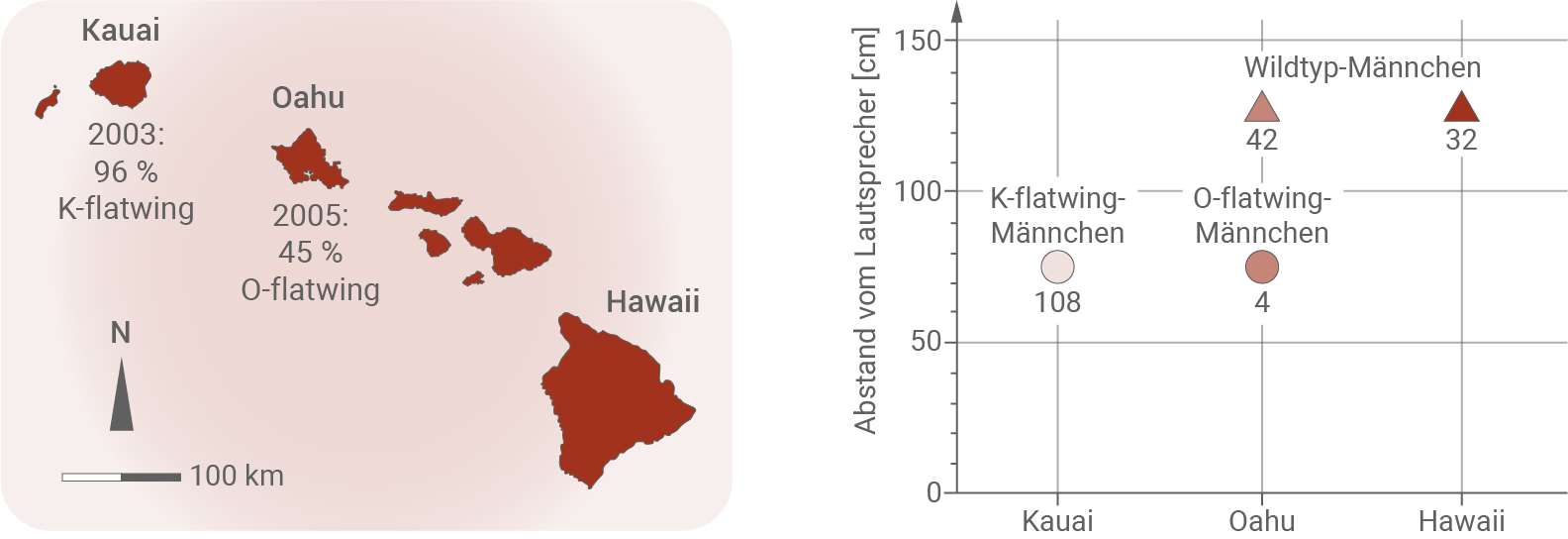
Abb. 2: Herkunft der untersuchten T. oceanicus-Populationen. Der jeweilige Anteil von flatwing-Männchen ist angegeben.
Abb. 3: Durchschnittlicher Abstand der T. oceanicus-Männchen vom Lautsprecher. Die Zahlen unter den Symbolen geben die Anzahl der getesteten Individuen an.
Material C: Vergleichende Untersuchungen der Flügelmorphologie
Auf Oahu und Kauai wurde eine Vielzahl von T. oceanicus-Männchen gefangen, um ihre rechten Vorderflügel zu untersuchen. Aus den dabei gewonnenen Messwerten wurden für jedes Männchen zwei Form-Maßzahlen A und B gebildet, die charakteristische Merkmale der Flügelmorphologie zusammenfassen. Anhand der Maßzahlen-Paare kann die Morphologie von Flügeln verglichen werden. Je ähnlicher sich zwei Flügel sind, desto näher liegen ihre Diagrammpunkte beieinander. Je unterschiedlicher zwei verglichene Flügel sind, desto weiter auseinander liegen die Diagrammpunkte. Die Maßzahlen-Paare wurden für die Vorderflügel von Wildtyp-Männchen und flatwing-Männchen sowohl von Oahu als auch von Kauai ermittelt und als Diagrammpunkte in ein Koordinatensystem eingetragen (Abbildung 4).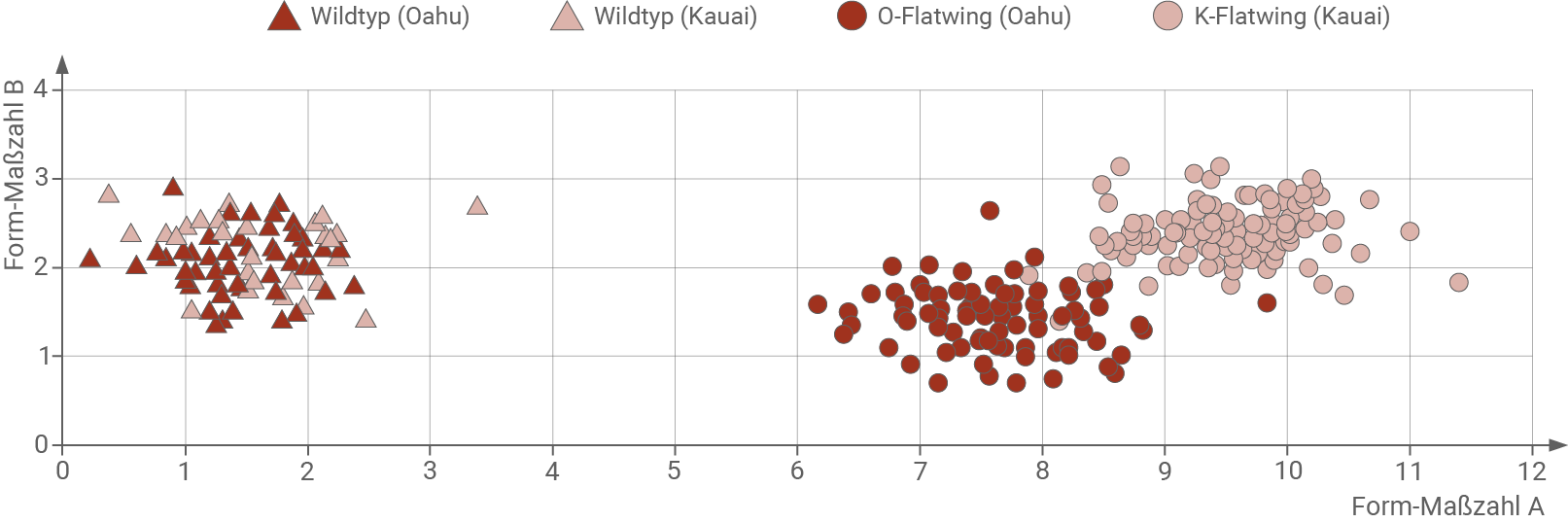
Abb. 4: Flügelmorphologie von T. oceanicus-Männchen. Jeder Diagrammpunkt entspricht dem Maßzahlen-Paar des Vorderflügels eines untersuchten Männchens.
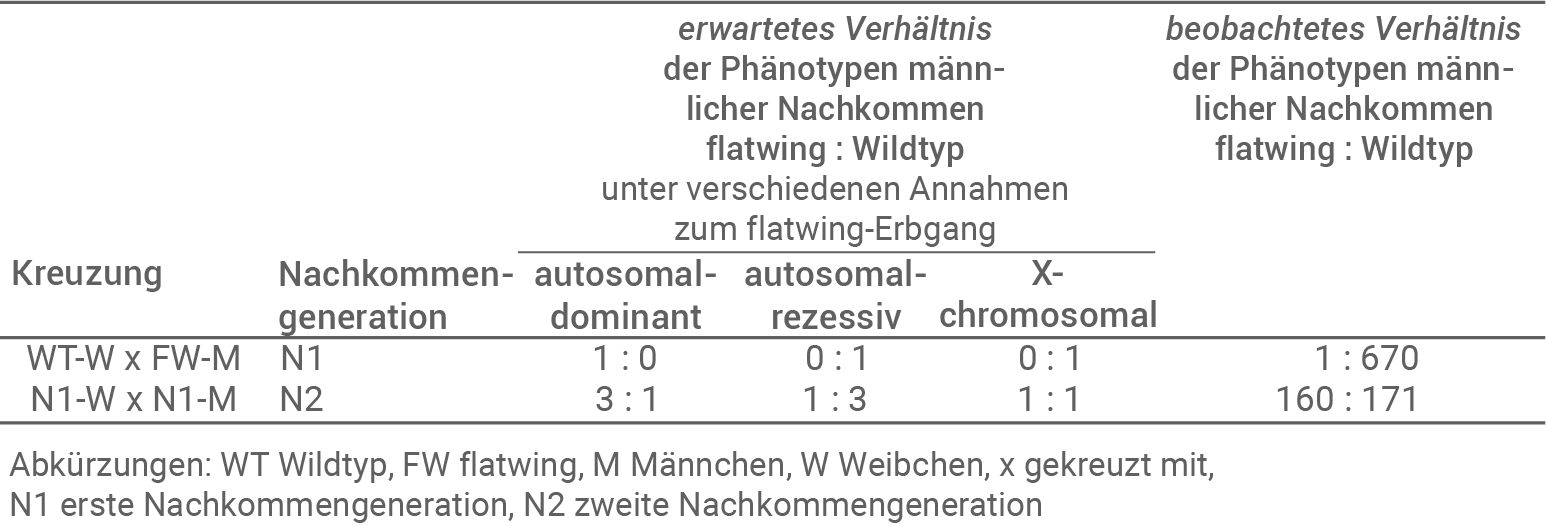
Material D: Genetische Grundlagen der flatwing-Morphologie
Um die genetischen Grundlagen der flatwing-Morphologie bei T. oceanicus zu untersuchen wurden Kreuzungsexperimente durchgeführt. Dazu wurden Weibchen von Kauai verwendet, die aus einer Wildtyp-Population entstammten und daher kein flatwing-Allel besaßen. Diese wurden mit Männchen aus einer stabilen flatwing-Population von Kauai gekreuzt. Die daraus entstandene erste Nachkommengeneration (N1) wurde dann untereinander gekreuzt, sodass man eine zweite Nachkommengeneration (N2) erhielt. Man bestimmte und zählte in N1 und N2 jeweils die Phänotypen der männlichen Nachkommen. Bei T. oceanicus erfolgt die Geschlechtsbestimmung über Geschlechtschromosomen, Weibchen besitzen zwei X-Chromosomen (XX) und Männchen eins (X0). Ein Y-Chromosom gibt es nicht. Tabelle 1 zeigt die erwarteten und die tatsächlichen Ergebnisse der Kreuzungen.
Tab. 1: Kreuzungsexperimente
Weiter lernen mit SchulLV-PLUS!
monatlich kündbarSchulLV-PLUS-Vorteile im ÜberblickDu hast bereits einen Account?
Hinweis: Die aufgeführten Lösungen sind stichpunktartig notiert. In der Prüfung ist ein die Stichpunkte inhaltlich umfassender Fließtext erforderlich, um die volle Punktzahl zu erreichen.
1.
Definition reproduktive Isolation:
- Unter reproduktiver Isolation versteht man verschiedene Aspekte, die zu einer dauerhaften Trennung des Genpools zweier Arten führen.
- Dabei unterbrechen Fortpflanzungsbarrieren den Genfluss zwischen Populationen, bis sich getrennte Genpools bilden, und die beiden Arten keine fruchtbaren Nachkommen mehr zeugen können.
- Die beiden Grillenarten unterscheiden sich durch ihren Lockgesang. Teleogryllus oceanicus stößt dabei einen hochfrequenten Gesang aus, während Teleogryllus commodus Gesang einer niedrigeren Frequenz erzeugt.
- Der Lockgesang innerhalb einer Art unterscheidet sich dabei kaum, und lockt damit artspezifisch paarungsbereite Weibchen an.
- Zwischen den beiden Grillenarten liegt somit eine ethnologische reproduktive Isolation vor, da die Isolation auf unterschiedliche Verhaltensweisen bezüglich des Lockgesangs zurückzuführen ist. Es handelt sich um einen präzygotischen Isolationsmechanismus.
- Im Gegensatz zu dem Lockgesang ist der Balzgesang innerhalb einer Art sehr variabel. Weibchen bevorzugen dabei diejenigen Männchen, die einen energieaufwändigen, hochfrequenten Balzgesang ausstoßen.
- Da Weibchen den energieaufwändigen Balzgesang bevorzugen, wird die reproduktive Fitness jener Männchen erhöht, die einen Balzgesang besonders hoher Frequenz ausstoßen. Das Weibchen kann daraus schließen, dass dieses Männchen über sogenannte „good genes“ verfügt, die es an seine Nachkommen weitergeben, und dadurch auch deren reproduktive Fitness erhöhen wird. Die reproduktive Fitness des Weibchens wird dadurch ebenfalls erhöht.
- Somit dient der Balzgesang der intersexuellen Selektion.
2.
Zusammenfassung der in den Abbildungen 2 und 3 dargestellten Inhalten:
- Im Jahr 2003 wurde auf Kauai eine größere Population Teleogryllus oceanicus mit verkürzter Schrillader vorgefunden. Diese stummen Individuen machten im selben Jahr schon ein Anteil von 96 % der Gesamtpopulation aus. Auch die Population auf Oahu wies 2005 schon einen Anteil von 45 % stummen Tieren auf.
- Die stummen Arten hielten in einem Experiment mit rund 75 cm einen deutlich geringeren Abstand zu Lockgesang-ausstoßenden Lautsprechern, als die Wildtyp-Art mit über einem Meter.
- Rufende Männchen haben den Selektionsnachteil, dass sie durch ihre Rufe die parasitäre Fliege anlocken, deren Larven das Männchen töten. Gleichzeitig locken sie paarungsbereite Weibchen mit den Rufen an, was wiederum einen Selektionsvorteil darstellt.
- Stumme Männchen haben insofern einen Selektionsvorteil, als dass sie O. ochracea nicht gezielt durch ihre Rufe anlocken. Sie werden dadurch nicht parasitiert, was ihre Überlebenschancen deutlich erhöht.
- Stumme Männchen können aber auch nur durch Zufall von paarungsbereiten Weibchen gefunden werden. Dieser Selektionsnachteil kann durch die räumliche Nähe zu rufenden Männchen ausgeglichen werden, da dadurch die Wahrscheinlichkeit erhöht wird, von Weibchen gefunden zu werden.
- Bei der divergenten Entwicklung entstehen ausgehend von einem gemeinsamen Ursprung morphologisch unterschiedlich ausgeprägte Varianten homologer Strukturen, da unterschiedliche Selektionsdrücke wirken.
- Konvergenter Entwicklung liegt ein gleicher Selektionsdruck zugrunde, der auf die beiden Arten wirkt. Die Entwicklung geht nicht auf einen gemeinsamen Ursprung zurück.
- Im Diagramm stimmt die Flügelmorphologie der beiden Wildtyp-Arten aus Kauai und Oahu weitestgehend überein.
- Die Diagrammpunkte der Wildtyp-Arten bilden im Vergleich zu den beiden flatwing-Arten eine deutlich getrennte Gruppe. Die Flügelmorphologie der Wildtyp- und flawting-Arten unterscheiden sich demzufolge stark.
- Auffällig ist außerdem die Trennung der Diagrammpunkte der beiden flatwing-Gruppen. Somit müssen sich auch deren Flügel morphologisch unterscheiden.
- Es ist zu erwarten, dass die Flügelmorphologie der Wildtyp-Arten sehr ähnlich ist, da auf den beiden Inseln vermutlich ähnliche Selektionsdrücke herrschen.
- Stumme Männchen könnten von Kauai nach Oahu verschleppt worden sein. Demzufolge wäre ihre Flügelmorphologie auch recht ähnlich. Diese Theorie ist aber eher unwahrscheinlich, da sich die Flügelmorphologie von K-flatwing und O-flatwing unterscheidet.
- Aufgrund der ähnlichen Selektionsdrücke auf Kauai und Oahu könnte die veränderte Flügelmorphologie auch unabhängig voneinander entstanden sein. Diese Hypothese wird durch die unterschiedlichen Maßzahlen der beiden Populationen und das zeitversetzte Auftreten der veränderten Flügel gestützt.
- Daher ist es sehr wahrscheinlich, dass es sich um eine konvergente Entwicklung der flatwing-Männchen auf Kauai und Oahu handelt.
- Alternativ könnte die unterschiedliche Flügelmorphologie der flatwing-Arten auch auf den Gründereffekt zurückzuführen sein.
3.
Zu erwartende Zahlenverhältnisse für die angegebenen Erbgänge:
- Autosomal-dominante Vererbung:
Bei diesem Vererbungsgang müssten beim Kreuzen von reinerbigen Wildtyp-Weibchen mit reinerbigen flatwing-Männchen nur Träger des flatwing-Gens in der ersten Nachkommengeneration entstehen. In der zweiten Nachkommengeneration sollte sich dann ein 3:1 Verhältnis zwischen Flatwing und Wildtyp einstellen. - Autosomal-rezessiv Vererbung:
Nimmt man diesen Vererbungsgang an, so sollten in der ersten Nachkommengeneration nur Wildtyp-Nachkommen entstehen. In der zweiten Nachkommengeneration sollte sich dann ein 1:3 Verhältnis zwischen Flatwing und Wildtyp einstellen. - X-chromosomale Vererbung:
Bei diesem Erbgang müssen alle Nachkommen der ersten Generation Wildtyp-Phänotypen aufweisen, da alle Individuen von dem homozygoten Weibchen ein X-Chromosom erhalten. In der N2 ist ein 1:1 Verhältnis zu erwarten, da die Männchen hemizygot sind, und mit einer Wahrscheinlichkeit von 50 % ein X-Chromosom des flatwing-Trägers oder des Wildtyp-Trägers erhalten.
- Es konnte gezeigt werden, dass es sich um einen X-chromosomalen Erbgang handelt, da die Zahlen dem zu erwarteten Aufspaltungsverhältnis für diesen Erbgang entsprechen.
- Ein Männchen in der N1 zeigte einen flatwing-Phänotyp, der bei einem X-chromosomalen Erbgang eigentlich nicht zu erwarten wäre. Dies könnte auf eine spontane Mutation auf dem X-Chromosom zurückzuführen sein.
4.
Entwicklung der stummen Grillen auf Kauai:
- Durch eine zufällige Mutation des Gens, deren Genprodukte für die Ausprägung der gezähnten Schrillader verantwortlich sind, entstanden Individuen mit veränderte Schrillader. Diese Individuen waren fortan nicht mehr in der Lage, den Lockgesang zu erzeugen.
- Diese Individuen brachten ihre Gene in den Genpool der Population ein, da sie im Gegensatz zu den singenden Männchen nicht parasitiert wurden, was ihre reproduktive Fitness erhöhte.
- Einige Teleogryllus oceanicus Weibchen müssen tolerant gegenüber dem veränderten Balzverhalten gewesen sein, und sich dennoch mit den Männchen gepaart haben.
- Bei einer starken Dezimierung der Wildtyp-Männchen könnten diejenigen Weibchen einen Selektionsvorteil haben, die tolerant gegenüber verändertem Balzverhalten sind. So konnten die Allele der flatwing-Männchen vermehrt in den Genpool der Population eingebracht werden.
- Es hat sich innerhalb einer sehr kurzen Zeit von zwei Jahren eine stabile Population von stummen flatwing-Männchen auf Kauai entwickelt.
- Aufgrund des Selektionsdrucks hin zu der mutierten Flügelvariante konnten die flatwing-Männchen eine höhere reproduktive Fitness erzielen, als die singenden Männchen.
- Im Jahr 2001 wurde eine deutliche Dezimierung der Population auf Kauai festgestellt. Der daraus resultierende Flaschenhalseffekt könnte durch Gendrift die Verbreitung des mutierten Allels begünstigt haben.
- Die Verbreitung dieser Allele konnte durch ihre Lokalisation auf dem X-Chromosom weiter beschleunigt werden.
- Da die Männchen von Teleogryllus oceanicus hemizygot sind, wirkt sich bei ihnen der Erhalt eines mutierten X-Chromosoms unmittelbar auf den Phänotyp aus. So können vorteilhafte Allele sehr schnell in einer Population ausbreiten.