Vorschlag A: Störche
Hinweis: Von den Vorschlägen A und B soll in der Prüfung nur einer bearbeitet werden.
Futtersuche der Störche
Werden Wiesen frisch gemäht, erscheinen dort oft Störche neben den Traktoren, um nach Schnecken, Mäusen und Fröschen zu suchen. Bisher war nicht bekannt, welche Sinnesorgane die Störche nutzen, um diese reichhaltige Futterquelle ausfindig zu machen. Dieser Frage sind Forschende nachgegangen.Nahrungsbeziehungen und Nahrungssuche
1
Gib jeweils eine Definition für die Fachbegriffe Trophieebene, Produzenten, Konsumenten und Destruenten an. Begründe, warum Nahrungsketten selten über vier Trophieebenen hinausgehen.
(12 BE)
2
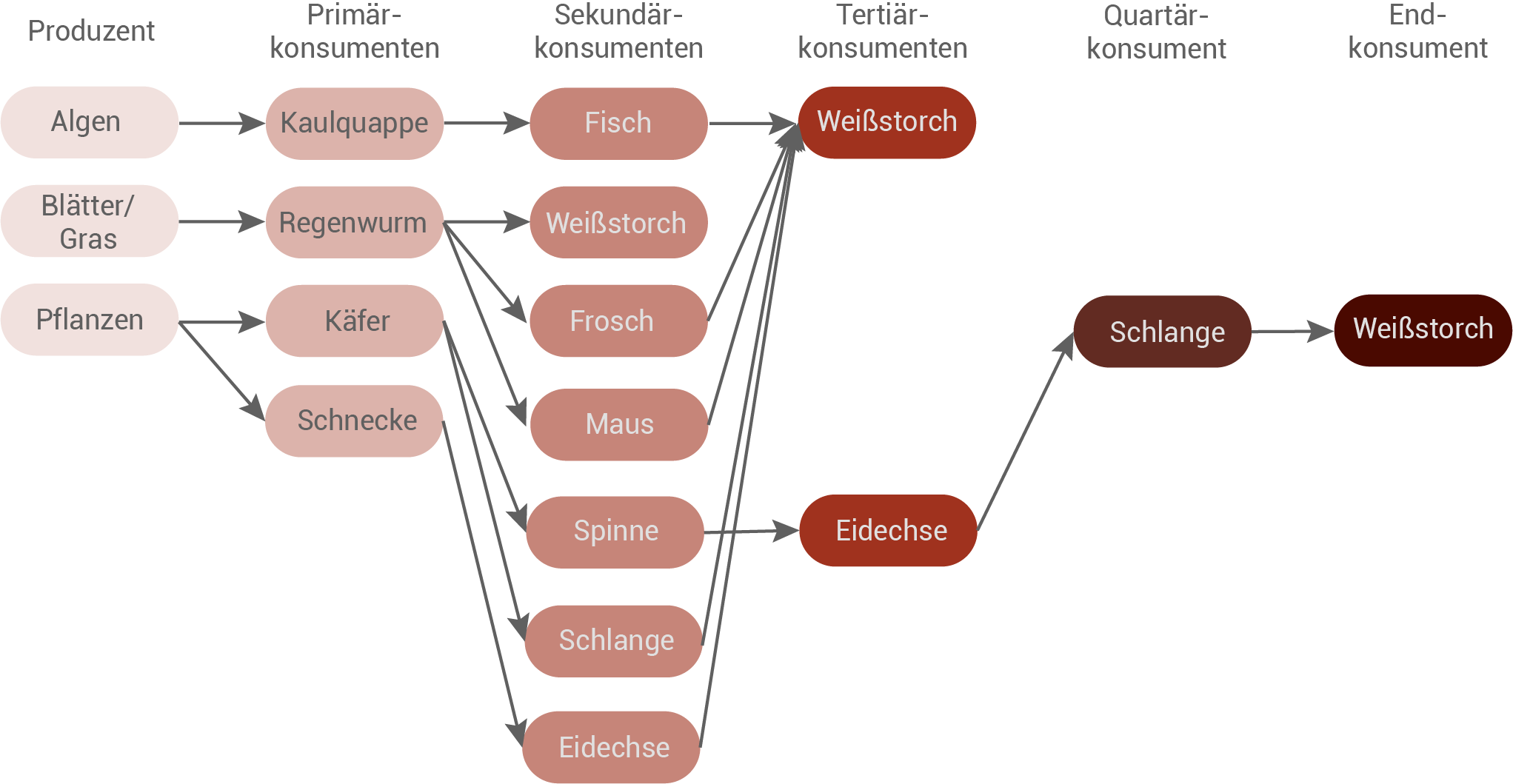
Entwirf aus den Angaben in Material 2 ein Nahrungsnetz und gib zwei Beispiele an, die zeigen, dass Organismen mehreren Trophieebenen zugeordnet werden können. (Material 1 und 2)
(10 BE)
3
Werte die Einzeluntersuchungen in Material 3 im Zusammenhang miteinander aus. (Material 1 und 3)
(18 BE)
Molekulare Grundlagen der Sinneswahrnehmung
4
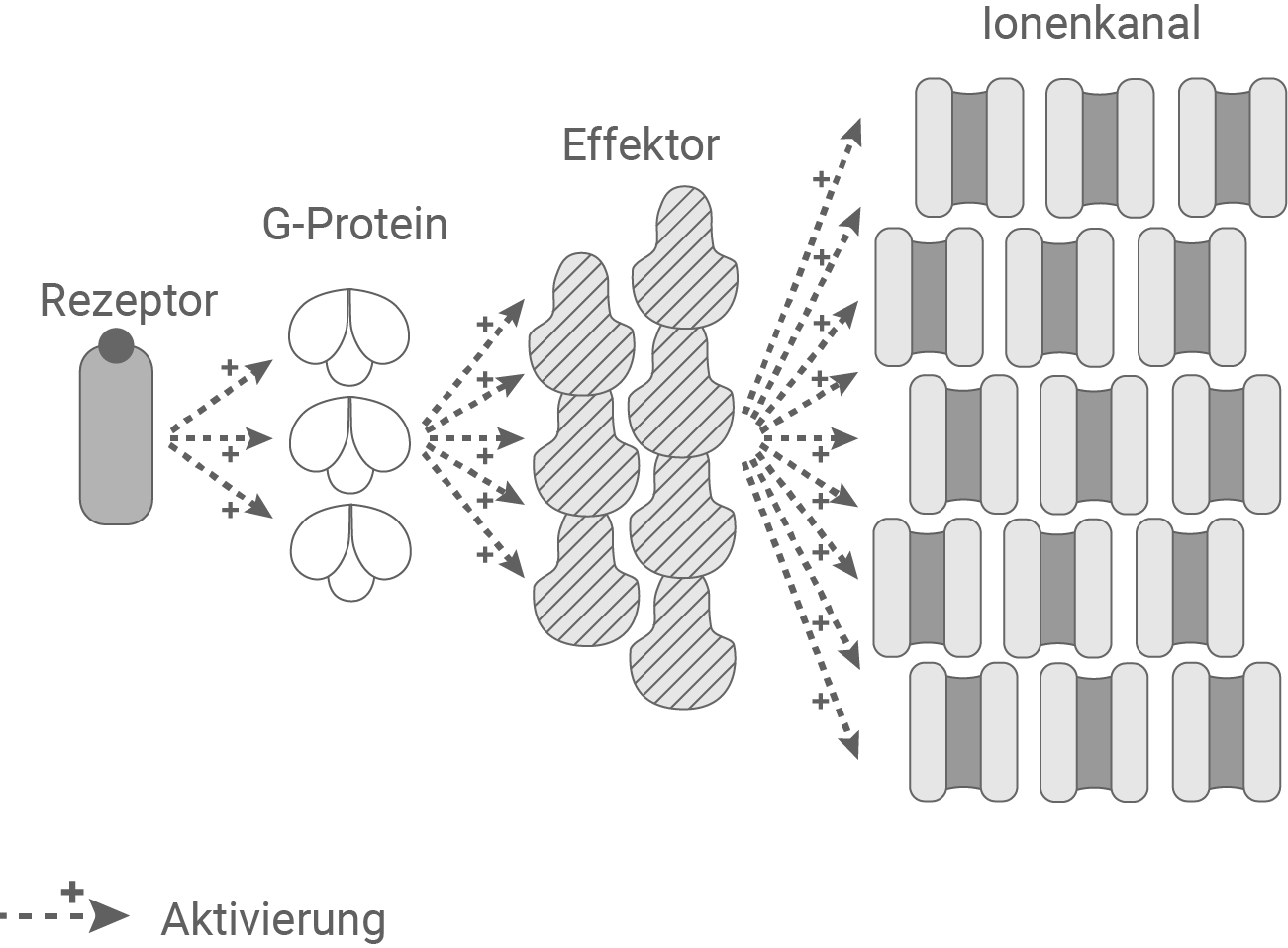
Beschreibe und vergleiche die in der Abbildung 4.1 dargestellten Prinzipien der Reizaufnahme und Signaltransduktion. Erkläre unter Berücksichtigung von Abbildung 4.2 den möglichen Vorteil der komplexeren Signaltransduktion in Zellen des Typs B im Vergleich zu Zellen des Typs A. (Material 4)
(8 BE)
5
Beschreibe und erläutere die Signaltransduktion in einem Stäbchen der Netzhaut bei Lichteinfall. (Material 5)
(10 BE)
6
Beschreibe und erläutere die Signaltransduktion in einer Geruchssinneszelle und vergleiche diese tabellarisch mit der Signaltransduktion in einem Stäbchen. (Material 5 und 6)
(22 BE)
7
Erläutere die Modellvorstellung zur Wahrnehmung unterschiedlicher Geruchsqualitäten anhand der Erregung durch einzelne Duftmoleküle (Abbildung 7.1 und 7.2). Stelle gemäß dem Modell die Stärke der Reaktion der verschiedenen Rezeptoren 1 bis 9 auf die Duftmoleküle F bzw. H dar. (Material 7)
(12 BE)
8
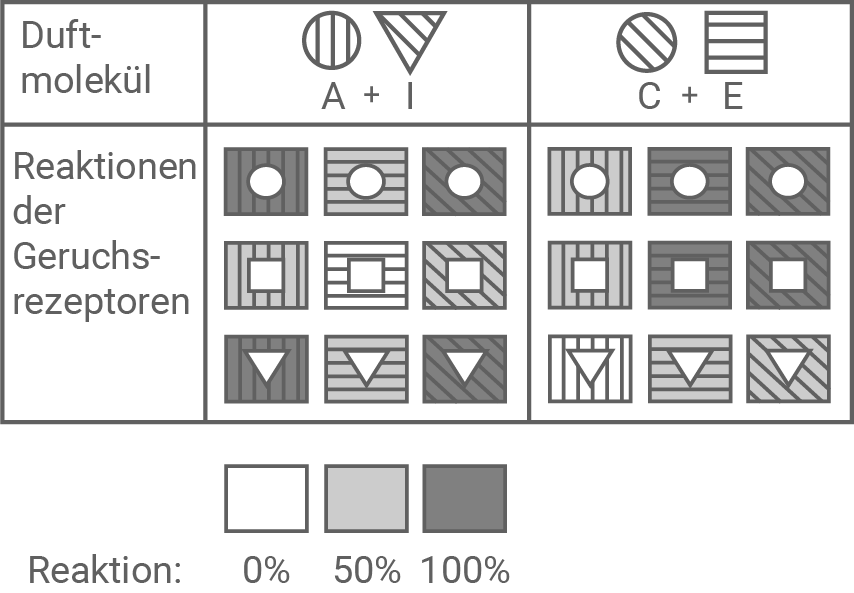
Stelle die Stärke der Reaktion der verschiedenen Rezeptoren auf das Gemisch der Duftmoleküle B + G entsprechend der in Abbildung 7.3 dargestellten Beispiele dar. Zeige, dass das Modell geeignet ist zu erklären, dass viel mehr verschiedene Geruchsqualitäten wahrgenommen werden können als unterschiedliche Rezeptoren vorhanden sind. (Material 7)
(8 BE)
Weiter lernen mit SchulLV-PLUS!
monatlich kündbarSchulLV-PLUS-Vorteile im ÜberblickDu hast bereits einen Account?Material 1
Der Weißstorch
Der Weißstorch (Ciconia ciconia) lebt in feuchten Gebieten in der Nähe von Seen oder Flüssen und ernährt sich von vielfältigen Kleintieren wie z. B. von Insekten, Schnecken, Fröschen und kleinen Nagetieren, die in hohen Wiesen Unterschlupf finden. Werden solche Wiesen gemäht, sind diese Tiere besonders leicht zu erreichen.

Material 2
Nahrungsketten des Weißstorchs
- Algen → Kaulquappe (Frosch) → Fisch → Storch
- Blätter/Gras → Regenwurm → Storch
- Blätter/Gras → Regenwurm → Frosch → Storch
- Blätter/Gras → Regenwurm → Maus → Storch
- Pflanzen → Käfer → Spinne → Eidechse → Schlange → Storch
- Pflanzen → Schnecke → Eidechse → Storch
- Pflanzen → Käfer → Schlange → Storch
Material 3
Nahrungssuche und Orientierung der Störche
Das Finden von Nahrung ist für Tiere überlebenswichtig. Über längere Distanzen können sie sich dabei verschiedener Sinne bedienen, z. B. des visuellen, akustischen oder olfaktorischen (Geruchssinn).
Ein europäisches Forschungsteam untersuchte die Orientierung von Weißstörchen bei der Futtersuche. Die Störche sammeln sich oft auf Wiesen, die gerade gemäht werden.
Das Forschungsteam beobachtete die aus ca. 70 Individuen bestehende Weißstorch-Population rund um die Stadt Böhringen am Bodensee sowohl aus Kleinflugzeugen heraus als auch mittels GPS-Ortung und Bodenbeobachtungen. Die Reaktion der Störche wurde jeweils an verschiedenen Tagen und an mehreren verschiedenen Orten beobachtet und dabei unterschiedlich viele Störche bzw. Storchengruppen untersucht.

Abbildung 3.1: Untersuchungsgebiet mit Beobachtungsflächen und Standorten der Störche
Dargestellt ist die Luftansicht des Untersuchungsgebiets mit einer Auswahl von Beobachtungsflächen sowie exemplarische Standorte der Störche und deren mögliche Anflugrouten.
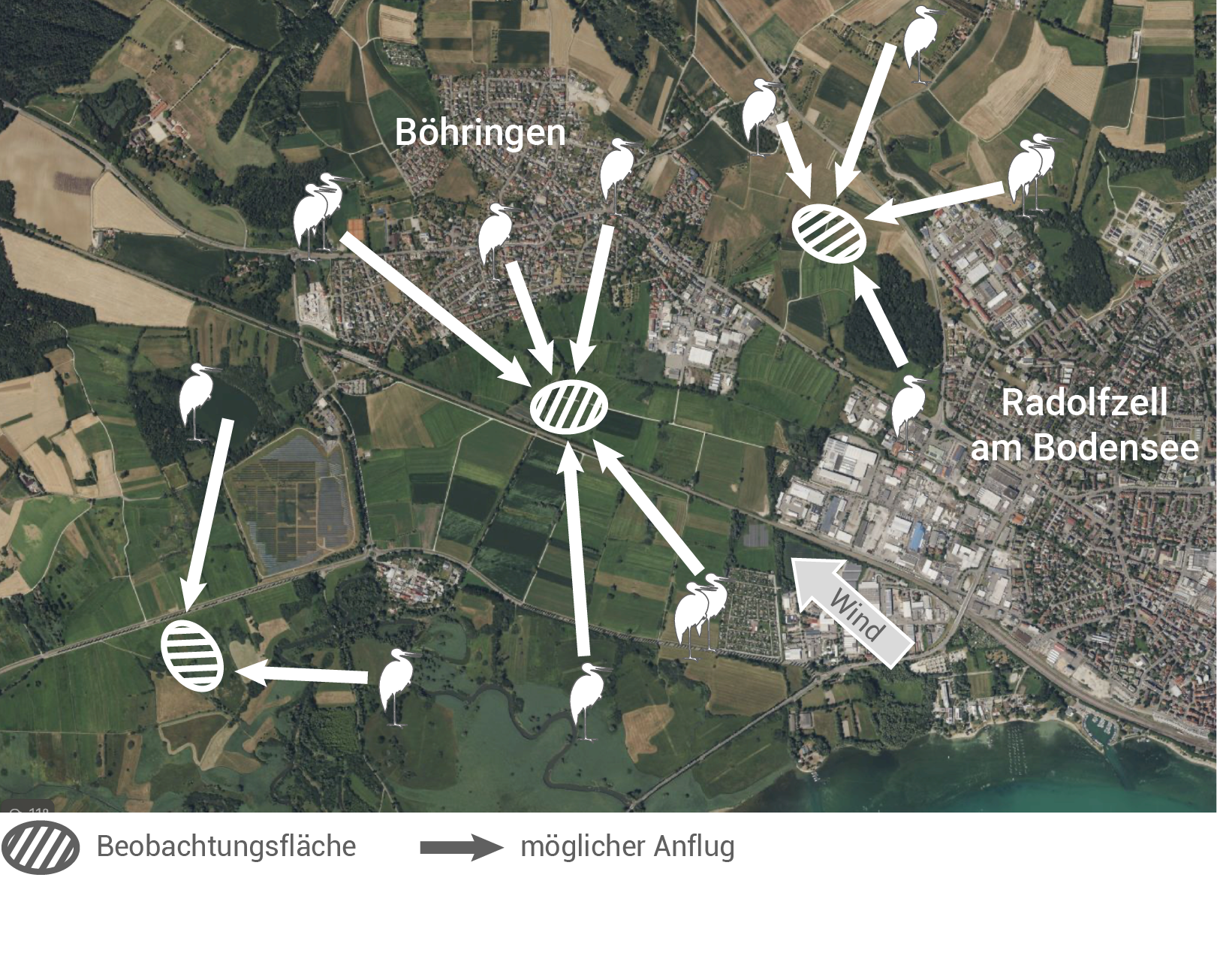
Abbildung 3.2: Reaktion der Störche auf frisch gemähte Wiesen bzw. Gras
Die Reaktion der Störche auf frisch gemähte Wiesen (A) wurde direkt während des Mähens beobachtet. Eine weitere Fläche (B) war bereits zu einem früheren Zeitpunkt gemäht worden. Auf dieser Fläche wurde frisch gemähtes Gras von einer 15 km entfernten Wiese verteilt.
Die einzelnen Symbole stellen entweder einzelne Individuen dar oder kleine Gruppen, deren Mitglieder optischen Kontakt miteinander hatten, also entweder nahe beieinander auf dem Feld standen oder gemeinsam flogen. Die Grad-Angaben zeigen den Winkel des Standorts bzw. des Anflugs der Störche auf die Untersuchungsflächen in Relation zur Windrichtung. 0° entspricht dabei der Richtung, aus der der Wind weht, 180° der Gegenrichtung.
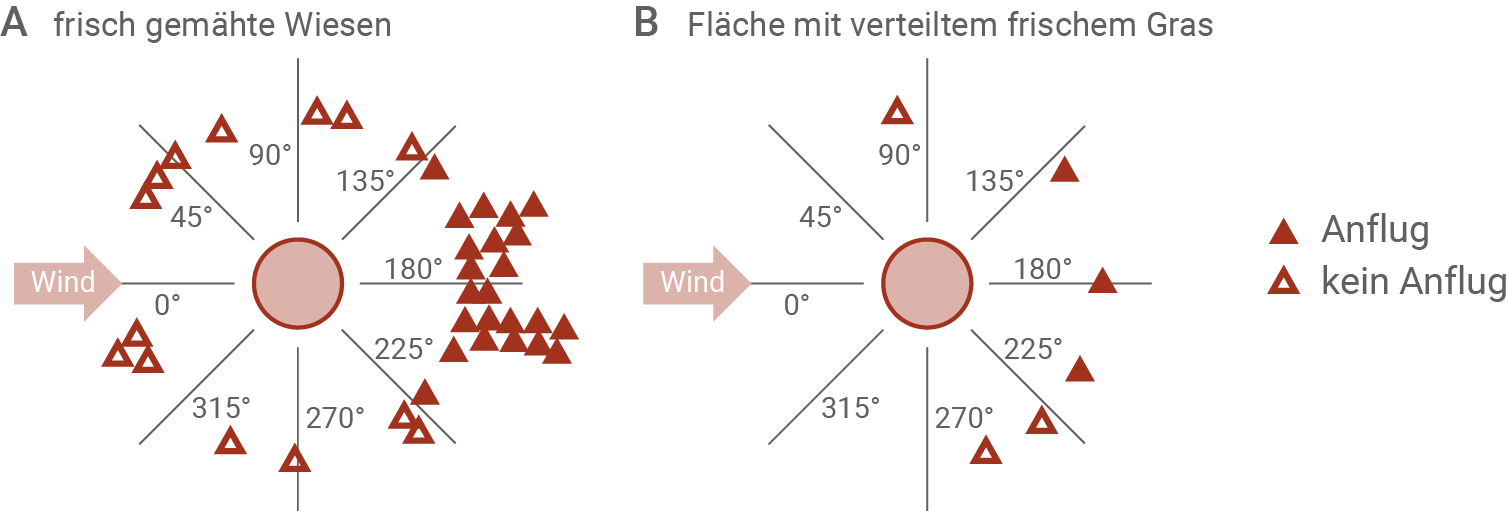
Abbildung 3.3: Reaktion der Störche auf Blattduftstoffe
Das Duftstoffgemisch, das den typischen Geruch frisch geschnittenen Grases erzeugt, enthält hauptsächlich drei verschiedene Substanzen.
Ein derartiges Gemisch wurde aus 20 m Höhe aus einem Ultraleichtflugzeug über eine ungemähte Wiese gesprüht (A). Alternativ wurde dieser experimentelle Ansatz mit reinem Wasser durchgeführt (B).
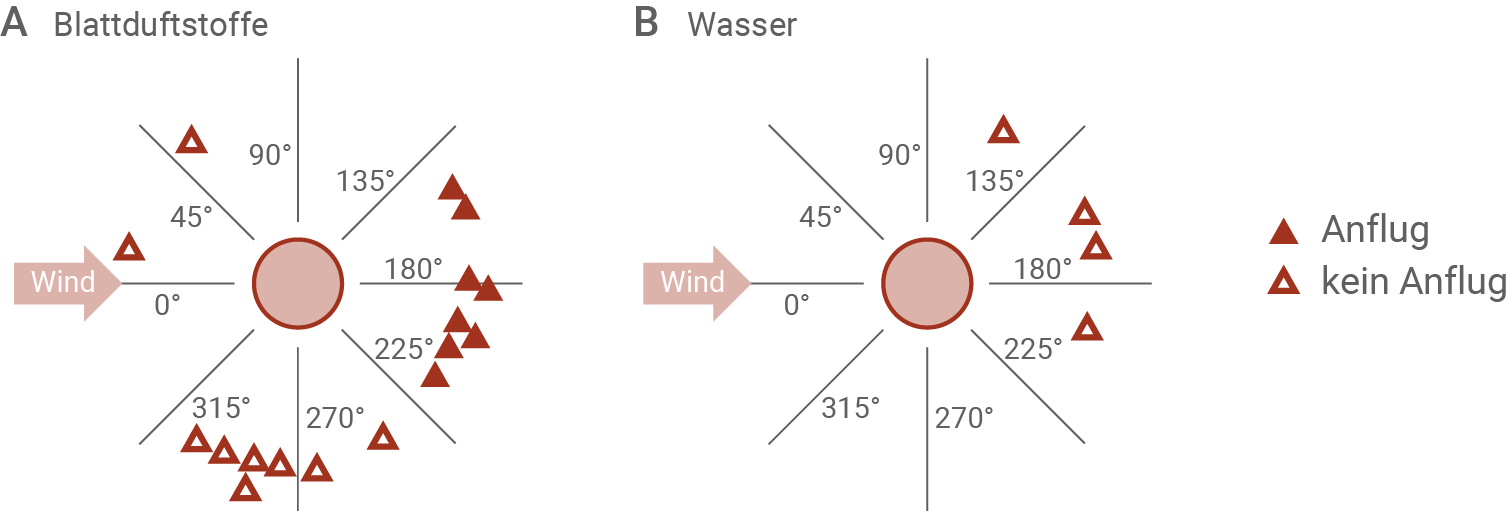
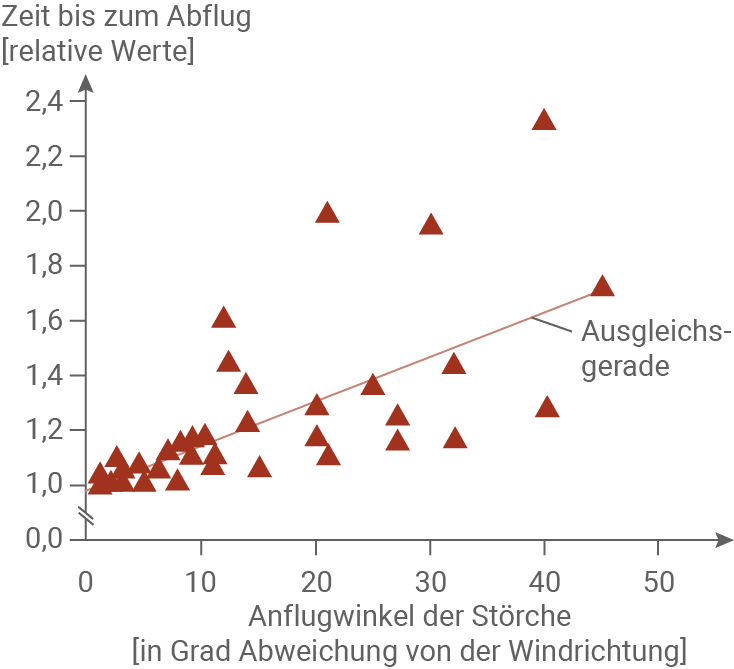
Abbildung 3.4: Reaktionszeit der Störche in Abhängigkeit vom Standort relativ zur Windrichtung
Es wurde ermittelt, wie schnell die Störche reagieren, wenn der Duft gemähten Grases an ihrem Standort angekommen ist. Die Reaktionszeit wurde in Bezug auf die Windgeschwindigkeit und Entfernung standardisiert, woraus sich relative Werte ohne Einheit ergeben. Es wurde kein Unterschied festgestellt zwischen der Beobachtung an frisch gemähten Wiesen, Flächen mit verteiltem frischem Gras und Besprühen mit Blattduftstoffen. In der folgenden Abbildung sind daher Ergebnisse aus allen drei Ansätzen zusammengefasst.
Material 4
Prinzipien der Reizaufnahme und Signaltransduktion in Sinneszellen
Abbildung 4.1: Rezeptortypen
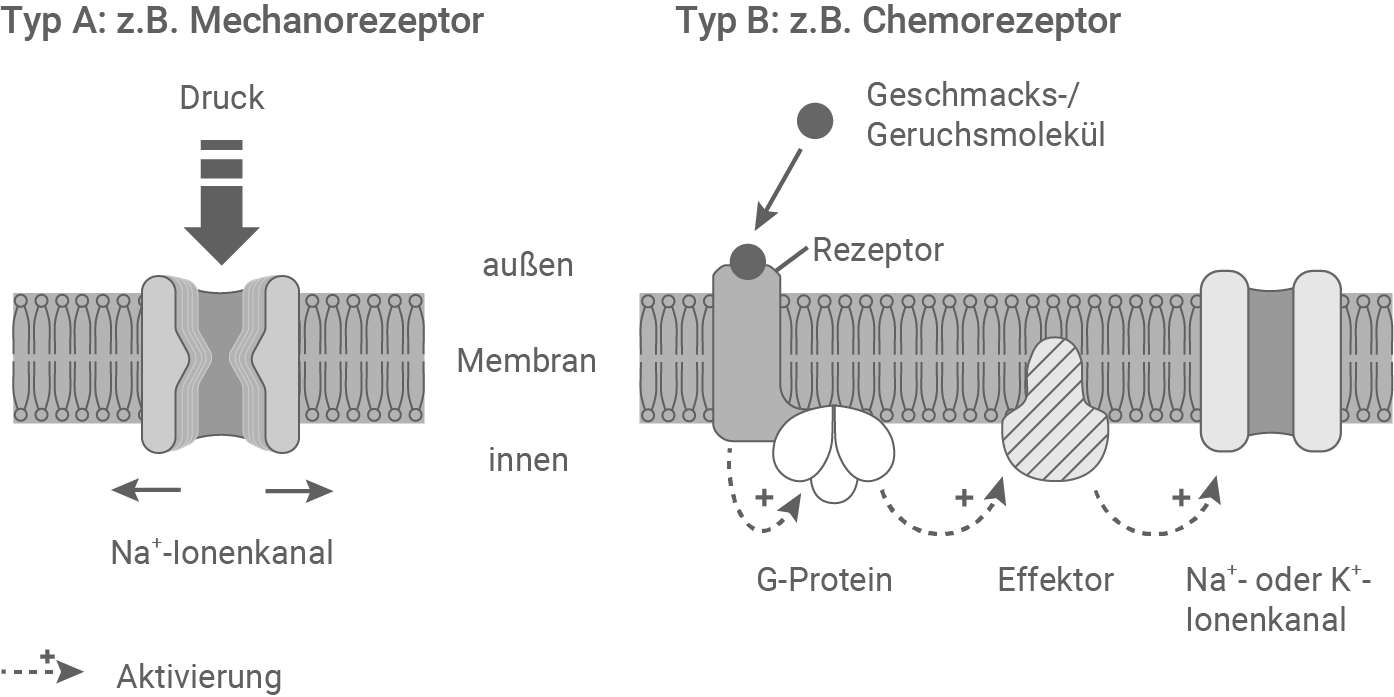
Abbildung 4.2: Signaltransduktion in Rezeptor-Zellen des Typs B
Material 5
Signaltransduktion in einer Lichtsinneszelle (Stäbchen)
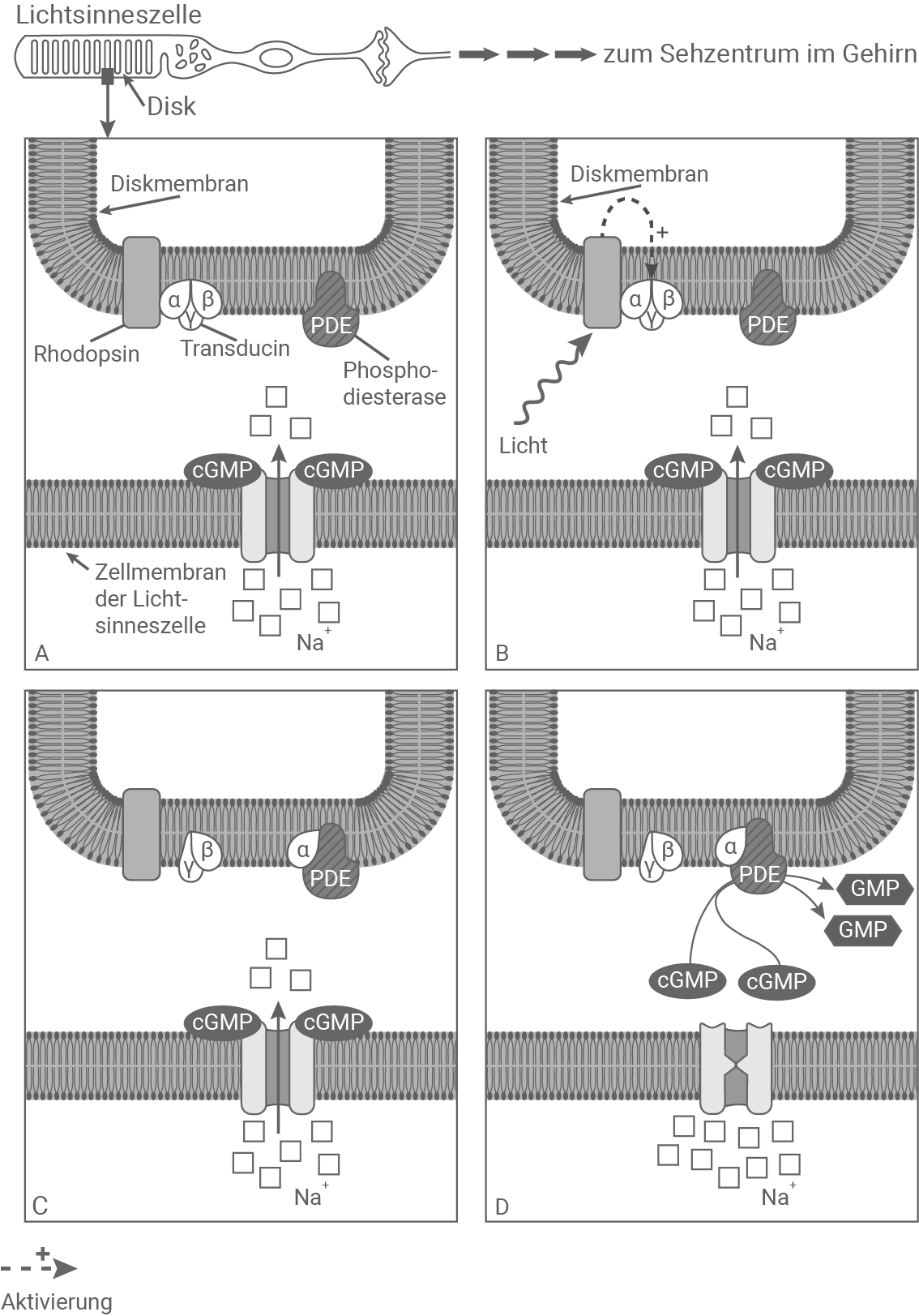
Material 6
Signaltransduktion in einer Geruchssinneszelle
Die anatomische und funktionelle Organisation des Geruchssinns von Vögeln ähnelt der aller anderen Wirbeltiere. Die Geruchssinneszellen sitzen am oder im Schnabel und ragen mit feinen Sinneshaaren (Cilien) in den Luftraum. An diesen Cilien wird Luft vorbeigeleitet. Einzelne darin enthaltene chemische Komponenten werden von spezifischen Rezeptormolekülen in den Cilien-Membranen erkannt, in neuronale Signale umgewandelt und diese an die entsprechenden Riechzentren (Riechkolben) im Gehirn weitergeleitet.
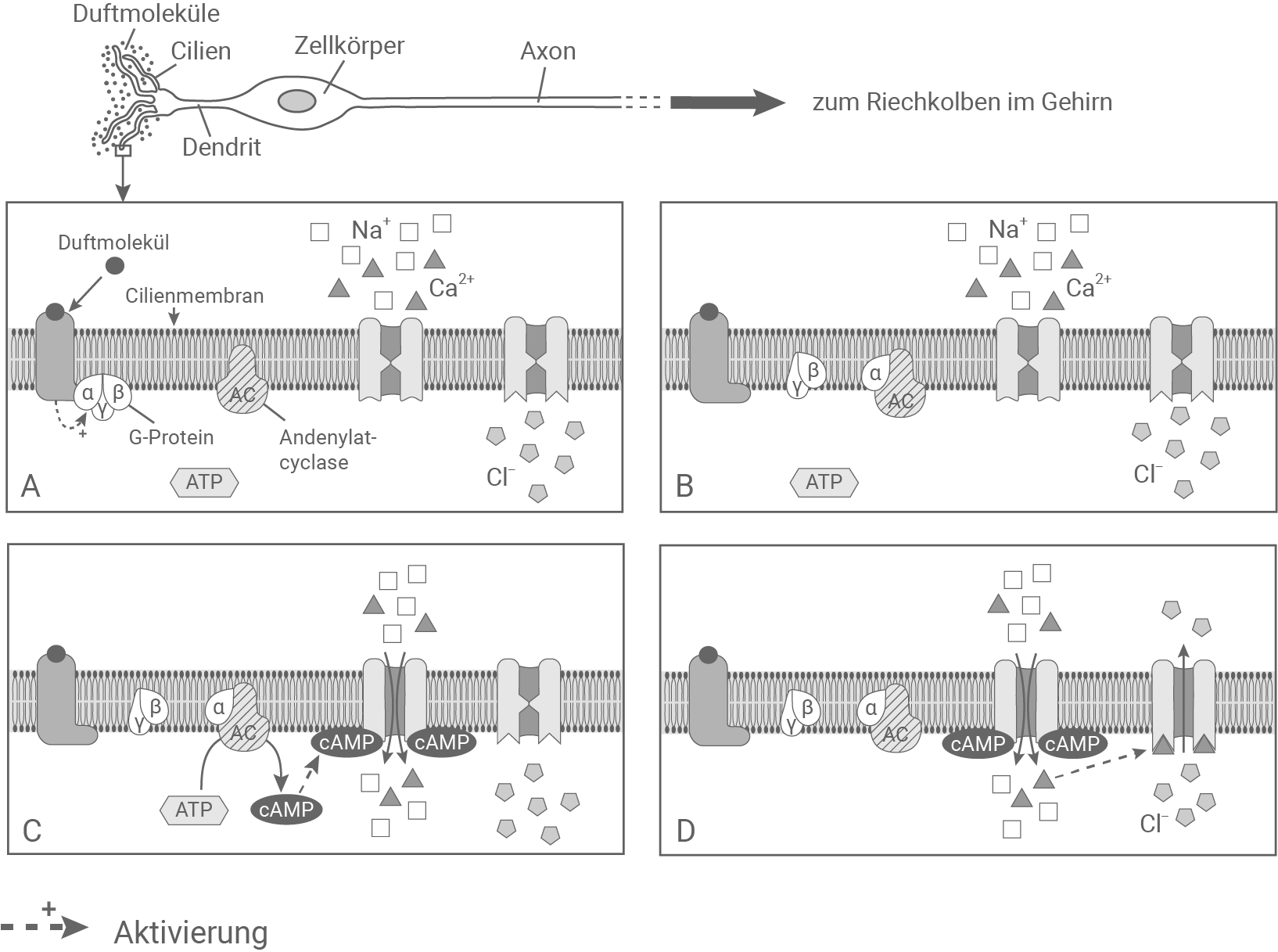
Material 7
Modellvorstellung zur Wahrnehmung unterschiedlicher Geruchsqualitäten
Bisher hat man bei Wirbeltieren ca. 1.000 verschiedene Riechsinneszellen identifiziert. Jede dieser Riechsinneszellen enthält in der Membran ihrer Cilien nur einen einzigen Geruchsrezeptortyp. Allerdings kann der Organismus ein Vielfaches an Geruchsqualitäten unterscheiden.
Das im Folgenden abgebildete Schema stellt ein mögliches Modell dar, das die Entstehung der verschiedenen Geruchseindrücke erklären soll. Geruchseindrücke entstehen durch die Kombination verschiedener Duftmoleküle. Es wird davon ausgegangen, dass jedes Duftmolekül mehrere charakteristische Eigenschaften hat. Im Modell sind beispielhaft zwei Eigenschaften durch Form und Schraffur dargestellt. Die Erregungen der verschiedenen Sinneszellen werden an das Riechzentrum im Gehirn weitergeleitet und dort verrechnet.
Abbildung 7.1: Verschiedene Duftmoleküle (A – I) und Geruchsrezeptoren (1 – 9)
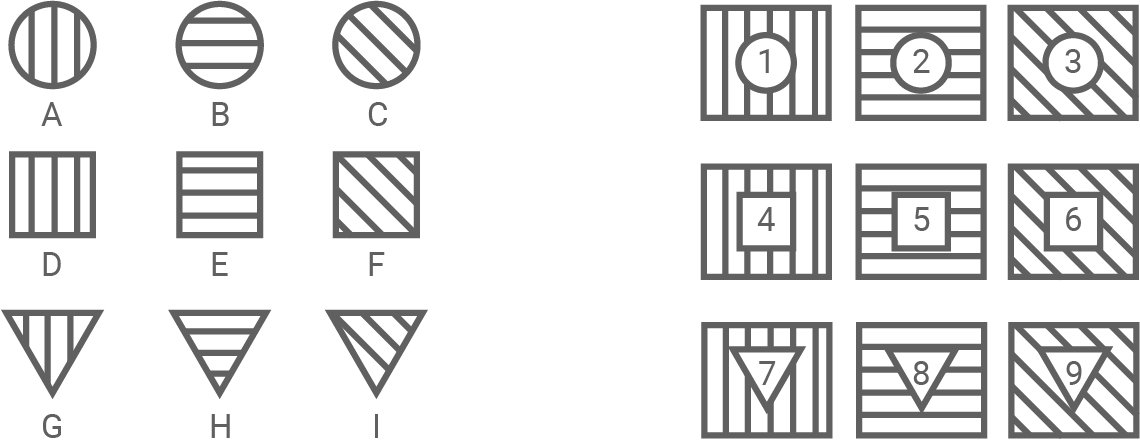
Abbildung 7.2: Reaktion der Geruchsrezeptoren 1 bis 9 auf verschiedene Duftmoleküle
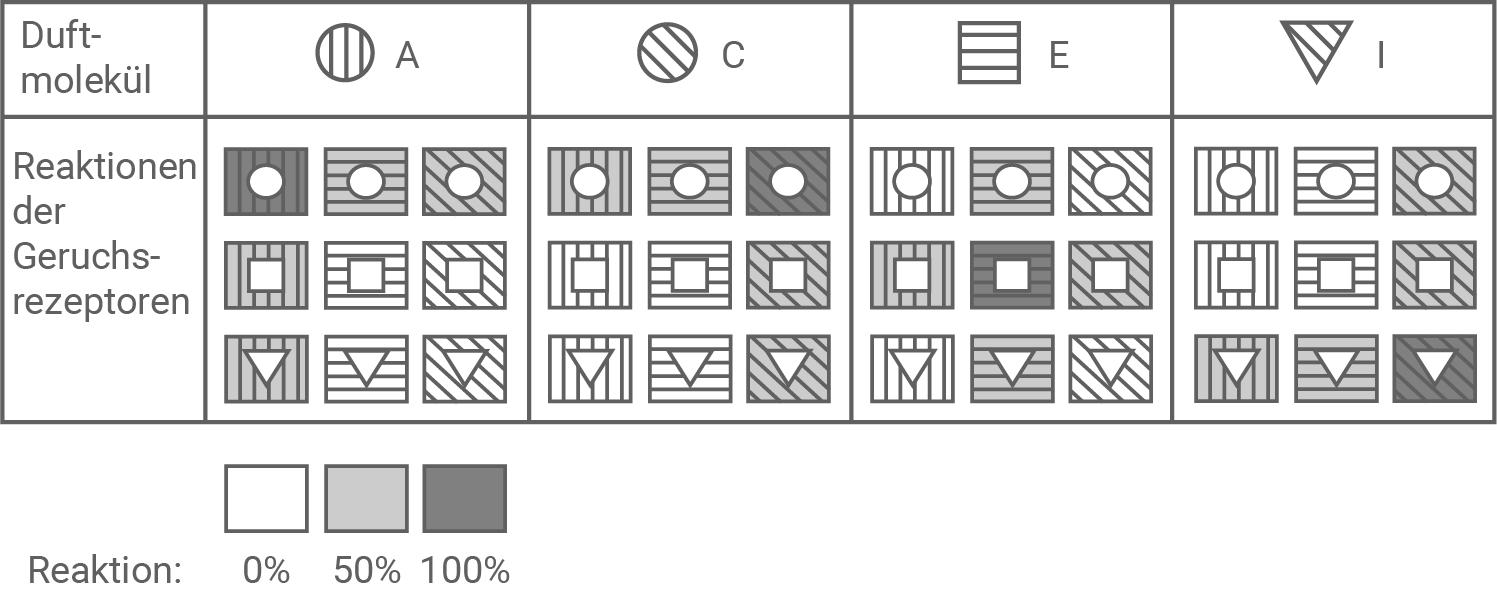
Abbildung 7.3: Reaktion der Geruchsrezeptoren 1 bis 9 auf verschiedene Gemische von Duftmolekülen
Weiter lernen mit SchulLV-PLUS!
monatlich kündbarSchulLV-PLUS-Vorteile im ÜberblickDu hast bereits einen Account?
1
Definitionen für Trophieebenen:
- Produzenten: Autotrophe Organismen, die durch Prozesse wie Photosynthese oder Chemosynthese organische Stoffe aus anorganischen Substanzen herstellen. Beispiele hierfür sind Pflanzen, Algen und Cyanobakterien. Sie bilden die Grundlage der Nahrungskette, da sie die primäre Energiequelle für alle Konsumenten sind. Ohne Produzenten wäre ein Energiefluss in Ökosystemen nicht möglich, da sie Sonnenenergie oder chemische Energie in verwertbare Biomasse umwandeln.
- Konsumenten: Heterotrophe Organismen, die organische Stoffe konsumieren, indem sie andere Lebewesen fressen. Sie werden in Primärkonsumenten (Pflanzenfresser wie Kühe), Sekundärkonsumenten (Fleischfresser wie Füchse) und Tertiärkonsumenten (Spitzenräuber wie Haie) unterteilt. Konsumenten sorgen für die Weitergabe von Energie in der Nahrungskette und regulieren gleichzeitig die Populationen anderer Organismen.
- Destruenten: Organismen wie Pilze und Bakterien, die abgestorbenes organisches Material abbauen und in anorganische Stoffe umwandeln. Dadurch schließen sie den Stoffkreislauf und machen Nährstoffe für Produzenten wieder verfügbar. Ihre Rolle ist essenziell, da ohne Destruenten Nährstoffe nicht recycelt würden, was langfristig zum Zusammenbruch eines Ökosystems führen könnte.
- Trophieebenen: Hierarchische Stufen in einer Nahrungskette, die die Position eines Organismus in Bezug auf die Energie- und Nährstoffflüsse beschreiben. Produzenten befinden sich auf der ersten Ebene, Konsumenten folgen in den höheren Ebenen. Destruenten sind nicht direkt Teil dieser Hierarchie, da sie auf alle Trophieebenen wirken.
- Mit jeder Trophieebene nimmt die verfügbare Energie ab, da nur etwa 10 % der Energie an die nächste Ebene weitergegeben werden. Die restliche Energie geht durch Stoffwechselprozesse und Wärmeverlust verloren. Diese Energieverluste machen es energetisch ineffizient, viele Trophieebenen zu unterhalten.
- Die abnehmende Energieübertragung begrenzt die Anzahl der Konsumenten auf höheren Ebenen, da die verfügbare Biomasse nicht ausreicht, um größere Populationen aufrechtzuerhalten. Spitzenprädatoren wie Haie oder Adler haben daher geringe Populationsdichten, was ihre Anfälligkeit gegenüber Umweltveränderungen erhöht.
2
Nahrungsnetz des Weißstorches:
 Beispiele für Organismen auf mehreren Trophieebenen:
Beispiele für Organismen auf mehreren Trophieebenen:
- Weißstorch: Der Storch kann mehreren Trophieebenen zugeordnet werden. Entscheidend ist dabei, welche Beute er wählt. Frisst er Fische, so ist er Sekundärkonsument. Frisst er Schlangen, die Eidechsen gefressen haben, so ist er Endkonsument. Frisst er Schlangen, die sich von Käfer ernährt haben, so ist er Tertiärkonsument.
- Eidechse: Ernährt sie sich von Schnecken gehört sie zu den Sekundärkonsumenten. Ihre Rolle im Nahrungsnetz ist flexibel und hängt von ihrer Beute ab. Frisst die Eidechse Spinnen, so gilt sie als Tertiärkonsument.
3
Auswertung der Untersuchungen zu Störchen:
Bei der Beobachtung der Nahrungssuche der Störche wurden Gruppen von je ca. 70 Tieren beobachtet. Da bei der Beobachtung darauf geachtet wurde, dass die Tiere keinen Sichtkontakt zu anderen Vögeln, oder der Wiesenfläche hatten, kann eine Verfälschung der Daten ausgeschlossen werden, da sich die Störche nicht an anderen Vögeln orientieren oder den Mähvorgang bzw. das Ausbringen von Gras oder Duftstoffen unmittelbar beobachten konnten.
Abbildung 3.1 zeigt das Untersuchungsgebiet von oben, und stellt mögliche Anflugrouten der Störche zu den Beobachtungsflächen dar. Dabei ist zu erkennen, dass die Tiere potenziell aus verschiedenen Richtungen den Beobachtungsplatz anfliegen können.
In Abbildung 3.2 ist die Reaktion der Störche auf frisch gemähte Wiesen bzw. Flächen mit verteiltem frischem Gras zu sehen. Dabei ist ein deutlicher Trend zu erkennen. Trifft der Wind die Störche in einem Bereich zwischen 135° und 225°, so wird der Beobachtungsplatz angeflogen. Ist der Winkel zu klein oder zu groß, dann wird der Platz nicht angeflogen. Dabei scheinen sich mehr Tiere in der Nähe der frisch gemähten Wiese zu befinden, als in der Nähe der Fläche, auf der das frische Gras verteilt wurde. Möglicherweise könnte der akustische Reiz des Mähvorgangs mehr Tiere angezogen haben, als wenn nur der Geruch des Grases als Reiz vorhanden war.
Aus Abbildung 3.3 geht hervor, dass die Blattduftstoffe des frisch gemähten Grases tatsächlich für den Anflug der Störche verantwortlich waren. Der Vergleich mit der Kontrolle mit Wasser zeigt, dass die Tiere nicht auf diese Kontrolle mit einem Anflug reagierten, aber einen Anflug starteten, wenn Blattduftstoffe ausgebracht wurden.
Die Grundlage für Abbildung 3.4 war eine Beobachtung der Zeit bis zum Abflug der Störche relativ zum Anflugwinkel. Die Abbildung zeigt, dass die benötigte Zeit mit zunehmender Abweichung von der Windrichtung steigt. Während in einem Abweichungsbereich bis zu etwa 15° die Streuung noch sehr gering ist, nimmt diese mit größeren Abweichungen von der Windrichtung zu. Das bedeutet, dass mit zunehmender Abweichung von der Windrichtung, die Zeit bis zum Abflug je nach Individuum stärker schwankt, als es bei kleinen Abweichungen der Fall ist.
Interpretation der Ergebnisse:
Zusammengefasst kann gesagt werden, die die beim Mähen einer Wiese entstehenden Blattduftstoffe den entscheidenden Reiz für den Anflug darstellen. Bei der Wahrnehmung des Duftstoffes spielt aber auch die Windrichtung eine entscheidende Rolle. Je geringer die Abweichung entgegen der Windrichtung, desto wahrscheinlicher und schneller ist ein Anflug. Neben dem olfaktorischen Reiz, kann auch der akustische Reiz, der beim Mähen einer Wiese entsteht, die Tiere anlocken.
4
Vergleich der Prinzipien von Typ A und Typ B Rezeptoren:
- Typ A (Mechanorezeptoren): Diese Rezeptoren reagieren auf physikalische Reize wie Druck oder Dehnung. Mechanische Veränderungen der Zellmembran führen zur direkten Öffnung spezifischer Ionenkanäle, wodurch Na+-Ionen in die Zelle einströmen und eine Depolarisation erzeugt wird. Dieser Prozess benötigt keine Zwischenschritte, was Mechanorezeptoren ideal für schnelle Reaktionen macht, wie etwa bei der Berührung oder dem Hören. Der Vorteil dieses Mechanismus liegt in der Geschwindigkeit, da die Signalweiterleitung ohne komplexe Kaskaden erfolgt.
- Typ B (Chemorezeptoren): Chemische Moleküle wie Duftstoffe binden an spezifische Rezeptoren, die über ein G-Protein eine Signalkaskade aktivieren. Diese Kaskade führt zur Aktivierung eines Effektors durch ein G-Protein, das wiederum aktivierend auf Natrium- oder Kalium-Ionenkanäle wirkt und so eine Depolarisation auslöst. Die Signaltransduktion ist langsamer als bei Typ A, erlaubt jedoch eine präzisere Verarbeitung und Verstärkung schwacher Signale. Die Signaltransduktion bei Chemorezeptoren funktioniert über viele Zwischenschritte. Bei jedem Zwischenschritt wird das Signal verstärkt (Signalamplifikation).
Im Vergleich zu Typ A ermöglicht Typ B eine stärkere Signalamplifikation durch die G-Protein-vermittelte Kaskade. Dadurch können selbst schwache chemische Reize erkannt und verarbeitet werden. Da Duft- und Geschmacksstoffe oft nur in geringen Konzentrationen oder in einem Gemisch mit anderen Molekülen vorliegen, ist eine spezifische Erkennung über einen passenden Rezeptor, sowie eine Verstärkung des Signals, von Vorteil.
5
Signaltransduktion in Stäbchen der Netzhaut:
Im Zustand der Dunkelheit liegt das trimere G-Protein Transducin an Rhodopsin in der Diskmembran der Lichtsinneszellen vor. In der Zellmembran der Lichtsinneszelle befindet sich ein Natrium-Ionenkanal. Das an den Kanal gekoppelte cGMP sorgt dafür, dass der Kanal im Ruhezustand geöffnet ist, und Natrium-Ionen in die Zelle einströmen können. Trifft nun Licht auf die Lichtsinneszelle, so wird dieses von Rhodopsin absorbiert. Das führt dazu, dass Transducin durch Rhodopsin aktiviert wird. Transducin dissoziiert von Rhodopsin. Während die beta- und gamma-Einheit von Transducin an der Membran verbleiben, wird die alpha-Einheit abgespalten und an die ebenfalls in der Diskmembran befindliche Phosphodiesterase gekoppelt. Daraufhin wandelt die Phosphodiesterase cGMP in GMP um. Der Abzug von cGMP von dem Natrium-Ionenkanal bewirkt, dass sich dieser schließt. Der Natrium-Ioneneinstrom ist unterbunden. Durch diese Hyperpolarisation wird das physikalische Signal Licht in einen elektrischen Impuls umgewandelt, der anschließend ans Sehzentrum im Gehirn übermittelt wird.
6
Signaltransduktion in einer Geruchssinneszelle:
Bindet ein Duftmolekül aus der Luft an einen Rezeptor in der Cilienmembran, so wird ein Rezeptor-gekoppeltes trimeres G-Protein aktiviert. Durch die Aktivierung dissoziiert es vom Rezeptor, und die alpha-Einheit wird abgespalten, während die beta- und gamma-Einheit an der Membran verbleiben. Die alpha-Einheit bindet an die Adenylatcyclase, die sich ebenfalls in der Cilienmembran befindet. Dadurch wird das im Inneren der Cilie befindliche ATP von der Adenylatcyclase in cAMP umgewandelt. Dieses bindet einen in der Cilienmembran lokalisierten Ionen-Kanal, und sorgt dafür, dass dieser sich öffnet. So wird ein Einstrom von Natrium- und Calcium-Ionen in die Cilie ausgelöst. Calcium-Ionen binden den benachbarten Chlorid-Ionenkanal in der Cilienmembran. Dieser öffnet daraufhin und Chlorid-Ionen können aus der Cilie ausströmen. Das löst die Depolarisation der Zelle aus, und das Signal kann an den Riechkolben im Gehirn übermittelt werden.
Vergleich der Signaltransduktion einer Geruchs- und einer Lichtsinneszelle:
| Kriterium | Geruchssinneszelle | Lichtsinneszelle | |
|---|---|---|---|
| Gemeinsamkeiten |
|
||
| Unterschiede |
|
|
|
7
Modellvorstellung zur Wahrnehmung unterschiedlicher Geruchsqualitäten:
Ein Geruchsmolekül kann verschiedene Eigenschaften haben. Im Modell sind zwei Eigenschaften eines Duftmoleküls durch Form (rund, quadratisch oder dreieckig) und Schraffur (vertikal, horizontal oder diagonal) dargestellt. Die passenden Rezeptoren weisen eine Stelle auf, die genau auf eine Kombination aus Form und Schraffur passt (Abb. 7.1).
Aus Abbildung 7.2 geht hervor, dass der die Reaktion eines Geruchsrezeptors umso stärker ausfällt, je besser seine Eigenschaften zum Duftmolekül passen. Sind Rezeptor und Duftmolekül in Form und Schraffur passgenau, so wird zu 100 % eine Reaktion ausgelöst (in diesem Fall ist der Rezeptor dunkelgrau gefärbt). Ist entweder Form oder Schraffur identisch, so wird zu 50 % eine Reaktion ausgelöst (der Rezeptor ist hellgrau gefärbt). Passen weder Form noch Schraffur von Duftmolekül und Rezeptor zueinander, so wird keine Reaktion ausgelöst (0%, weißer Rezeptor).
Unterschiedliche Geruchsqualitäten können entstehen, da ein Duftmolekül sowohl den passgenausten als auch Rezeptoren, die in bestimmten Eigenschaften mit dem Duftmolekül übereinstimmen, binden kann. Im Gehirn werden die ankommenden Signale miteinander verrechnet, was schließlich zu einem bestimmten Eindruck von einem Geruch führt.
Reaktion der Rezeptoren 1-9 auf die Moleküle F und H:

8
Reaktion auf Duftmischungen:
 In einem Gemisch aus Duftmolekülen werden diejenigen Rezeptoren zu 100 % aktiviert, die sowohl zu einem der Duftstoffe passgenau sind, als auch diejenigen, die zu einer Kombination der Eigenschaften der Duftstoffe passen. So werden zum Beispiel auch die Rezeptoren für eine runde Form und diagonale Schraffur von Duftmolekül A+I aktiviert, obwohl diese Eigenschaften nicht bei A oder I allein zu finden sind.
In einem Duftgemisch sind verschiedene Duftmoleküle zu finden. In diesem Fall werden auch diejenigen Rezeptoren zu 100 % aktiviert, die zu einer Kombination der Eigenschaften der Duftmoleküle passen. Im Gehirn werden die ankommenden Signale miteinander verrechnet, was schließlich zu einem bestimmten Eindruck von einem Geruch führt. Dabei kann ein Duftmolekül allein einen anderen Eindruck auslösen, als wenn es in Kombination mit anderen Molekülen in einem Gemisch auftritt.
In einem Gemisch aus Duftmolekülen werden diejenigen Rezeptoren zu 100 % aktiviert, die sowohl zu einem der Duftstoffe passgenau sind, als auch diejenigen, die zu einer Kombination der Eigenschaften der Duftstoffe passen. So werden zum Beispiel auch die Rezeptoren für eine runde Form und diagonale Schraffur von Duftmolekül A+I aktiviert, obwohl diese Eigenschaften nicht bei A oder I allein zu finden sind.
In einem Duftgemisch sind verschiedene Duftmoleküle zu finden. In diesem Fall werden auch diejenigen Rezeptoren zu 100 % aktiviert, die zu einer Kombination der Eigenschaften der Duftmoleküle passen. Im Gehirn werden die ankommenden Signale miteinander verrechnet, was schließlich zu einem bestimmten Eindruck von einem Geruch führt. Dabei kann ein Duftmolekül allein einen anderen Eindruck auslösen, als wenn es in Kombination mit anderen Molekülen in einem Gemisch auftritt.